







Breeder Nutrition and Chick Quality
By Marcus Kenny and Carolyne Kemp, Aviagen - This article by Aviagen discusses how the nutrition of the breeder hen affects the chicks physiology at hatching. 11 July 2005
11 July 2005
 15 minute read
15 minute read

The developing embryo and the
hatched chick are completely
dependent for their growth and
development on nutrients deposited in
the egg. Consequently the physiological
status of the chick at hatching is greatly
influenced by the nutrition of the breeder
hen which will influence chick size,
vigour and the immune status of the
chick.
| Table 1. The necessary change in hatchery or broiler performance to equalise profitability when breeder feed cost is changed by 1% per tonne (for example from £UK 140.00/tonne to £UK141.40/tonne or £UK138.60/tonne). | |||
| Hatch of total eggs (%) | 0.24 | ||
| 42 day liveweight (g) | 7.4 | ||
| 42 day FCR | 0.0015 | ||
| 42 day mortality (%) | 0.07-0.45* | ||
| *depending on age of mortality. Calculated using input-output values for UK industry 2003 (Kemp and Kenny 2003). | |||
The financial effects
Nutritional decisions for breeders need
to take account of the overall economics
of the whole production cycle.
Table 1 shows the changes in hatchery
and broiler performance that are required
to equalise the effect of a 1%
increase in breeder feed cost on the profitability
of the whole production cycle.
Only one of these changes is required
to have the necessary economic effect;
in practice all are likely to move positively
making the measurements of any
one change difficult.
The calculations are done under typical
UK 2003 conditions and they show quite
clearly that small improvements in bird
performance are required to ‘pay’ for
more expensive breeder feed.
Conversely, apparent savings in
breeder feed cost can readily lead to an
overall loss if small changes in broiler
performances are ignored.
Similar economic analyses have been
conducted by Mississippi State University
which, based on US integration
2002 costs, demonstrates that a measurable
improvement in progeny liveability
as a result of hen diet change can be
profitable.
The key point is that trying to cut the
cost of a breeder feed may easily reduce
the profitability of the overall enterprise.
Influence of feed allocation
Underfeeding the hen can have an
impact on chick quality and this is particularly
noticeable in the early production
period. Modern hybrid parent flocks
commence production at a faster rate
than in the past and consequently egg
output increases over a shorter time span
during the early laying period. Feed allocations
during this period have not necessarily
increased in line with this egg
production trend. Low feed allocation
intake by young commercial breeder
flocks has been shown to compromise
nutrient transfer to the egg, resulting in
increased late embryonic death, poorer
chick viability and uniformity.
In a recent study by Leeson (2004)
broiler breeders were fed different levels
of feed through peak production varying
from 140 to 175 grams. Although the
increased feed allocation increased
bodyweight there was no influence on
egg size, however chick weight was
influenced by feed allocation (Table 2).
Of equal importance is the effect of
overfeeding on ovarian development. In
experimental studies ad libitum feeding
has been the most widely used model for
overfeeding which can result in excessive
follicular development or Erratic
Oviposition and Defective Egg Syndrome
(EODES).
Flocks with EODES generally have
poor shell quality, a reduced duration of
fertility and poor hatchability. It is also
known that fewer sperm will survive but
it is not clear how the surviving sperm
are affected and if they generate a
weaker embryo. The same authors also
warn that the effect of aggressive feeding
two to four weeks after photostimulation
reduces productive performance
throughout the life of the flock.
In this period the bird switches from
primarily growth to a reproductive state.
The young birds’ reproductive hormone
system is not mature enough to deal with
high nutrient intakes; nutrients are
instead metabolised to egg yolk lipid
which contributes to excess follicle
development.
Research shows that nutrient supply to
the broiler breeder is of consequence to
chick quality and production performance.
This places greater emphasis on
the nutritionist providing the correct
nutrient density diet and the flock manager
to provide appropriate feed intake
to the bird coming into lay.
| Table 2. The effects of breeder feed levels on chick weight. | |
| Peak breeder feed (g/b/d) | 30 week breeder chick weight (g) |
| 140 | 40.3 |
| 147 | 40.0 |
| 155 | 41.5 |
| 162 | 41.7 |
| 169 | 41.8 |
| 175 | 42.0 |
Diluted breeder diets
The use of diluted breeder diets is receiving a lot of attention in Western Europe on the basis of improvements in bird welfare. Experimental work feeding low energy density diets to young parent stock gave a delayed onset of oviduct development, increased early egg size, faster development of the embryo and a higher live weight of day old chicks. When broiler mortality was above average, low density broiler breeder feeds gave a significant reduction in mortality of offspring. Other experimental work showed improvements in breeder productive performance when diluted diets were fed in the rearing period.
Vitamins
Vitamins are involved in most metabolic
processes and are an integral part of
foetal development, therefore the consequence
of suboptimal levels of these
nutrients in commercial diets are known
to result in negative responses to both
parent and offspring performance.
Vitamins account for about 4% of the
cost of a breeder feed, so economising
on vitamin inclusion rates is rarely an option
| Table 3. Some practical recommendations for vitamin supplementation of breeder feeds (Fisher and Kemp 2001). |
|||
| Vitamin | Leeson & Summers (1997) | DSM | Ross |
| A (iu/g) | 7 | 10-14 | 13 |
| D3 (iu/g) | 3 | 2.5-3.0 | 3 |
| E (mg/kg) | 25 | 40-80 | 100 |
| K (mg/kg) | 3 | 2-4 | 5 |
| Thiamine (mg/kg) | 2.2 | 2-3 | 3 |
| Riboflavin (mg/kg) | 10 | 8-12 | 12 |
| Pyridoxine (mg/kg) | 2.5 | 4-6 | 6 |
| B12 (mg/kg) | 0.013 | 0.02-0.04 | 0.03 |
| Nicotinic acid (mg/kg) | 40 | 30-60 | 50 |
| D-pantothenic acid (mg/kg) | 14 | 12-15 | 12 |
| Biotin (mg/kg) | 0.2 | 0.2-0.4 | 0.3 |
| Folic acid (mg/kg) | 1 | 1.5-2.5 | 2 |
The levels of vitamin supplementation
recommended by different sources have
been summarised in Table 3. Generally
there is a shortage of information on vitamin
requirements of broiler breeders
especially when related to offspring performance.
Most of the breeder work is
quite dated and since that time breeder
performance has changed.
It would be impossible to review all the
literature in this article, however a
review of work on fat soluble vitamins,
biotin and pantothenic acid have shown
that vitamin E has the largest impact on
progeny.
| Table 4. Impact of dietary breeder vitamin status on bodyweight, enzyme activities, tissue characteristics and immunity of progeny. | ||
| Vitamin | Progeny response | |
| Vitamin A | Increased liver vitamin A in embryonic and chick liver but decreased vitamin E, carotenoids and ascorbic acid. Surai et al. (1998). | |
| Carotenoids | No positive impact on chick growth, organ development or humoral immunity in chicks five weeks post hatching. Haq et al. (1995). | |
| Carotenoids | Transferred from the hen to the yolk but not absorbed well by the embryo and subsequent chick. Haq and Bailey (1996). | |
| Carotenoids and Vitamin E | Carotene, vitamin E, and their combination improved and vitamin E lymphocyte proliferation, but only vitamin E improved humoral immunity. Haq et al. (1996). | |
| Vitamin E | Vitamin E levels of 150 and 450mg/kg increased passively transferred antibody levels in chicks to Brucella abortus up to seven days of age. Jackson et al. (1978). | |
| Vitamin E | Increased vitamin E in chicks’ yolk sac membrane, liver, brain and lung all of which had reduced susceptibility to peroxidation. Surai et al. (1999). | |
| Vitamin E | Increased progeny antibody titers to sheep red blood cells at hatch. Boa-Amponsem et al. (2001). | |
| Vitamin E and Selenium | Increased liver glutathione activity in chicks. Increasing selenium increased selenium dependant glutathione peroxidase in chick liver. Surai (2000). | |
| Vitamin D | Tibial calcium was increased at two weeks post hatching and tibial ash increased at four weeks of age by increased vitamin D3. Ameenudin et al. (1986). | |
| Vitamin K | Chicks from hens fed vitamin K deficient diet had reduced tibial glutamic acid levels at day one and 28 post hatching but tibial glutamic acid was restored by supplementing the chick diet with vitamin K. Lavelle et al. (1994). | |
| Biotin | Foot pad dermatitis and incidence of breast blisters were decreased in some trials in chicks from hens fed biotin fortified diet. Harms et al. (1976). | |
| Biotin | As biotin increased in the hens’ diet, yolk and chick plasma also increased. Biotin concentration in chick plasma was poorest from young hens. Whitehead (1984). | |
| Pantothenic acid | Liveability of chicks was best when hens were fed 20mg/kg diet of pantothenic acid. Utno and Klieste (1971). | |
| Adapted from M. Kidd 2002 | ||
The production and economic effects
of vitamin E supplementation are best
shown by Hossain et al (1998) where a
basal corn soya feed was supplemented
with 25, 50, 75 and 100mg/kg vitamin E.
The effects on hatchability were not
significant; however the best hatchability
was obtained at 50mg/kg at 52 weeks.
Offspring immune response continued
to increase up to 100mg/kg. In the same
studies higher final bodyweights at 42
days, improved FCR and reduced mortality
were observed in chicks from eggs
which had been injected with vitamin E
in ovo.
Haq et al., (1996) working with very
high levels of vitamin E (134mg/kg versus
412mg/kg) found no growth response
to 21 days and an improvement in FCR
for the offspring of hens receiving the
supplemental feed.
In other studies the combination of
selenium and vitamin E to broiler breeders
has been shown to increase liver glutathione
activity of progeny.
In general it seems to be justified to
supplement practical breeder feeds with
100mg/kg vitamin E.
There appear to be mixed reports on
the efficacy of vitamin C; some experiments
suggest a positive response, but a
more recent study failed to detect any
benefit on any production parameter.
This lengthy study used corn soya diets
supplemented with 75mg/kg stabilised
vitamin C which when analysed recovered
49mg/kg which might explain the
variability of response.
The influence of increased vitamin levels
fed to young parent stock on progeny
performance is an area which has
received significant commercial interest.
Work conducted at Aviagen Ltd has
shown chicks derived from 31 week old
parent stock fed elevated levels of vitamins
showed improved growth to 11
days and reduced mortality compared to
chicks derived from 42 and 45 week old
parents.
Similar responses have been found in
the field where chicks derived from
young parents fed increased levels of vitamins have benefited in terms of viability
and liveability. Perhaps this supports the
need for further work exploring the vitamin
requirements of the breeder in the
early production period.
Whitehead (1991) proposes that a basis
for making recommendations is to feed
vitamin levels that maximise the resulting
level in the egg.
For vitamins with active transport
mechanisms (thiamine, riboflavin, biotin,
cobalamin, retinol and cholecalciferol)
these levels reflect the saturation of binding
proteins.
Levels derived in this way include
10mg/kg for riboflavin and 250 microgram/
kg for biotin. Whitehead (1991)
contrasts this level of riboflavin with the
conventional requirement (4mg/kg in this
case) but the higher figure – the upper
limit to nutritionally useful range – may
be a better guide to good commercial
practice.
| Table 5. Blood cell count of the broiler derived from parents fed high or low vitamin and mineral levels (Rebel et al 2004). | |||
| Breeder low vitamins/minerals | Breeder high vitamins/minerals | ||
| Heterophil | 5.3 | 3.8 | |
| Lymphocyte | 4.6a | 21.4b | |
| Monocyte | 1.1 | 5.3 | |
| Basophil | 0.0a | 5.4b | |
| Table 6. Summary of minerals fed to breeders shown to have an effect on progeny performance. | ||||
| Growth | Liveability | Immune function | Skeletal | |
| Fluoride | X | |||
| Phosphorus | X | |||
| Selenium | X | |||
| Selenomethionine | X | X | ||
| Zinc | X | X | X | |
| Zinc and methionine | X | |||
| Adapted from M. Kidd 2002 | ||||
Vitamins and chick immunity
Reference has already been made to the
effect of vitamin E on chick health and
immune function, while other vitamins
have been researched none show the
same degree of effect as vitamin E.
Table 4 summarises work investigating
the effect of different vitamins fed to
breeders and consequent impact on
progeny health.
Recent work by Rebel et al (2004)
investigated the effects of several elevated
levels of vitamins and trace elements
fed to breeders and broilers on the
immune system of birds infected with
malabsorption syndrome.
Broilers derived from breeders fed elevated
vitamins and mineral levels had
increased numbers of leukocytes at day
old which indicated stimulation of the
immune system (see Table 5).
Fig. 1. The effect of protein-to-energy ratio in the breeder feed on chick weight at hatch (Spratt and Leeson 1987). 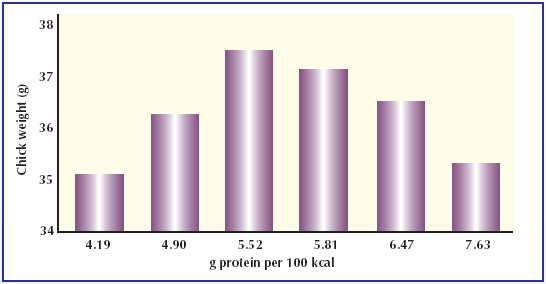 |
Major minerals
Calcium, phosphorus, sodium, potassium, magnesium and chloride are involved in shell formation hence general improvements in shell quality lead to better egg and chick quality. Variations in maternal phosphorus supply have been shown to influence bone ash of young but not older progeny. Broiler performance was not affected by these treatments so the practical significance of this work is not clear but the use of relatively low phosphorus levels in breeder diets, while benefiting egg shell quality, may not lead to the best possible bone integrity in the early stages of growth.
Trace minerals
Most interest in this field has centred on
the use of chelated minerals which have
been shown to increase deposition in the
egg and transfer to the tissues of the hen
and the embryo.
Most recent work has focused on the
antioxidant status of breeders, embryos,
offspring and the role of selenium.
Surai (2000) has shown the role of
Selenomethionine on both vitamin E and
glutathione peroxidase levels in eggs,
embryos and chicks up to 10 days of
age.
The economic benefits of using
Selenomethionine compared with
sodium selenite have been examined in
a number of unpublished field trials in
the UK. Hatchability improvements
ranged between 0.5-2.0 chicks per 100
eggs and in another trial 0.3-0.7 chicks
per 100 fertile eggs. Few of these tests
involve a proper assessment of subsequent
broiler performance although
comments about chick quality are generally
positive.
In one of the commercial trials mentioned
an improvement of 0.5% in mortality
and cull rate at 10 days was
observed when organic selenium
replaced sodium selenite.
Research has indicated that the
improvements in chick immunity as a
result of mineral fortification of hen diets
may result in improved liveability.
Flinchum et al. (1989) demonstrated
that leghorn breeders fed supplemental
zinc methionine to a zinc adequate diet
had progeny with improved survival to
an E. coli challenge. Similar improvements
to progeny liveability were seen
with breeders fed supplemental zinc and
manganese amino acid complexes.
Table 6 is a summary of those minerals
which, when fed to breeders, have an
effect on progeny performance.
Nutrient levels in the breeder diet
There is clear evidence that a high protein
to energy ratio depresses hatchability,
and probably chick performance.
The experiment by Whitehead et al.
(1985) shows the effect of excess protein
where the higher protein level reduced
reproductive performance, producing
3.1 fewer chicks per 100 fertile eggs.
Chick quality was also reduced so that
the difference in saleable chicks was 4.0
per 100 fertile eggs.
The effect of energy protein ratio in the
breeder feed is shown in Fig. 1. This
emphasises both the effects of excess
and inadequate protein, and also indicates
that the optimum level is quite
steeply defined.
According to this trial the optimum
protein level is at 5.52g protein per
100kcal which converts to an optimum
of 15.18% protein for a diet containing
2,750cal/kg of feed.
The protein level of the diet and its
ratio to energy is important not only for
parent performance but also for chick
quality.
| Table 7. Commercial comparison of breeder feeds based on wheat or maize (400g/kg). | ||
| Advantage of maize over wheat based feed | ||
| Mortality during lay (%) | -1.7 | |
| Total eggs (per hen housed) | +3.8 | |
| Hatching eggs (per hen housed) | +4.8 | |
| Hatching/total eggs (%) | +0.9 | |
| Hatch of set eggs (%) | +0.6 | |
| Hatch of fertile eggs (%) | +1.1 | |
| Second quality chicks | -0.1 | |
| Based on a comparison of two commercial houses each containing 6500 female grandparent breeders. Data to 58 weeks (Ross Breeders, unpublished data, 1998). | ||
The effect of feed ingredients
There is evidence of improved breeder
performance when maize is compared to
wheat as the main cereal in breeder
feeds. From a survey of many depleted
commercial flocks overall hatch of fertile
eggs in the UK based on wheat diets and
Brazil based on maize diets is 83.3 and
86.2 per 100 eggs respectively.
Other management factors may contribute
to this difference in hatchability
other than cereal source; male management
is very good in Brazil and the
resulting high fertility may also contribute
something to this difference.
Unpublished commercial development
trials from the Netherlands and Aviagen
Ltd grandparent flocks (see Table 7) support
this observation.
The most likely benefit of maize is
probably in shell quality and thickness.
From the same data average poorer
shells with specific gravity of <1.08
accounted for 26.1% of eggs from wheat
fed hens and 17.1% from maize fed.
Studies of hatching losses showed less
late dead embryos (>18 days) and less
bacterial contamination. These two
responses are expected with eggs of better
shell quality.
Evidence about fat levels and sources is
conflicting but there is no question that
this is an important consideration. Added
fat levels should be kept low in breeder
feed (1-3%) and preference given to
unsaturated vegetable oils rather than
saturated animal fats. Work from
Mississippi State University compared
maize oil and poultry fat and generally
supported the use of more unsaturated
fat (see Table 8).
Maize oil increased 21 day bodyweight
over that of poultry fat and improved
slaughter weight of broilers in comparison
to equal levels of poultry fat and
lard.
| Table 8. Experiments comparing fat sources and/or levels for broiler breeders. | ||
| Reference | Fats compared | |
| Brake (1990) | PF | |
| Brake et al. (1989) | PF | |
| Denbow & Hulet (1995) | SBO, PF, FO | |
| Peebles et al. (1999a, b) | CO, PF, LA | |
| Peebles et al. (2000a) | PF, CO, LA | |
| Peebles et al. (2000b) | PF, CO, LA | |
| Fats: PF – poultry fat; SBO – soybean oil; FO – fish oil; CO – corn oil; LA – lard | ||
Summary
Over and undersupply of nutrients into
and through lay can have a very significant
impact on breeder production and
quality of progeny. This places greater
emphasis on the nutritionist providing
the correct nutrient density diet and the
flock manager to provide appropriate
feed allocation in lay.
Addition of micronutrients to the
breeder has been shown to be beneficial
to progeny quality especially in the early
production period. Use of specific
dietary ingredients such as maize can
affect breeder performance and progeny
quality. Both on economic grounds and
on biological grounds, high quality nutrition
of breeders is well justified.
Source: Aviagen - June 2005


